
Laurent J.
Catoire
Research Director
Miri
Garrigues
pHD STUDENT
Claudia
Zilian
Post-doc
Nahya
Nguyen Armengaud
PHD STUDENT
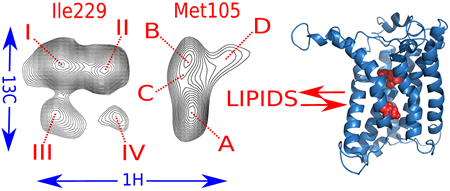




Thanks to a perdeuteration and a fine tuned biochemistry, we were able to obtained NMR data at a resolution never observed in the field. This provides direct evidence that BLT2 explores a complex landscape that includes four different conformational states for the unliganded receptor (Fig. 3). The relative distribution of the different states is modulated by ligands and the sterol content of the membrane, in parallel with the changes in the ability of the receptor to activate its cognate G protein. This demonstrates a conformational coupling between the agonist and the membrane environment that is likely to be fundamental for GPCR signaling.
Jean-Louis Banères
(IBMM, Montpellier)
(Marjorie Damian, Jacky Marie)
Eric Guittet
(ICSN, Gif/Yvette)
(Ewen Lescop, Nelly Morellet)